“Contours of Destruction”: How they work and why they are important for nature conservation

· 25 min read
I like to tell my colleagues about the so-called destruction contours. Normally, biogeocenosis is a homeostat. In response to various disturbances — logging, burning, grass from recreation, etc. — negative feedback is activated in the form of restorative successions (demutations). They restore the original living cover with its inherent mosaic, nutritional conditions, soils and humidity, etc., but only so far there are few “traces” of external influences in the form of different types of disturbances.
When their “network” passes a certain limit of density and/or intensity of disturbances, instead of the expected negative feedback, a positive one is triggered, changes occur that are outwardly similar to restorative successions, but do not eliminate the disturbances, but, on the contrary, strengthen them: spreading their space (their individual “spots” creep and merge with each other, like a tear in rotten fabric) and increasing the disturbance in each individual “spot.” In the extreme, this eliminates the previous community and establishes in its place a new, anthropogenically modified one.
The following describes the three most striking and “fresh” examples of this. The first is the clear collapse of hopes for “global greening”: planetary progress of vegetation, an increase in phytomass, restoration, and expansion of natural biomes responding to “carbon dioxide fertilization” — an exponential increase in its content in the atmosphere in the last 50 or 100 years due to anthropogenic causes, responsible for global warming. In the last 20–25 years, the opposite process has been reliably shown — an increase in the “burning” of carbon “stored” in peats, mortmass, and soil carbon during global warming, instead of the expected increase in its binding by vegetation or burial by swamps, precisely because the biomes of the planet are already 50 (or 100) years are too broken.
After it was reliably shown in 2015 that global warming exists, and that it is caused by human activity, the deniers of both changed their tunes and began to say that “warming will only get better”: the rise in carbon dioxide levels, going uncontrollably, stimulates the development of vegetation, the expansion of forests, the overgrowing of deserts, the growth of favorable climate instead of the growth of extremes expected by alarmists, in general, apple trees will bloom on Mars.
This “global greening” hypothesis was first proposed in the 2000s and has gained momentum in recent years.
It is known that in real high-quality disinformation there should be about 60% truth if for the public, or 95% if for specialists.
“Global greening” is closer to the second number, it has a rational grain that is important and interesting to us biologists: this is the hypothesis of “counteracting change”, reflecting the ability of the biosphere to regulate disturbance in the form of increased emissions of CO2 and CH4, as was the case in the pre-agricultural period (Friedman , Eremkin, 2015).
But, taken out of the context of the planet’s real biomes, of different types and, alas, on more than a third of the original area already eliminated (or severely disturbed) by man, this true thought becomes an error or misinformation (depending on the tenacity with which they insist on it despite the many facts that do not fit into this explanation, and of different types). Unlike the situation in the Pliocene, when comparable concentrations of greenhouse gases had purely positive consequences (the Sahara was mostly savanna, rivers flowed there, etc.), today’s exponential increase in the concentrations of the main greenhouse gases, CO2 and CH4, is by no means a separate natural process, but an integral part of the anthropogenic destruction of natural biomes and its consequences.
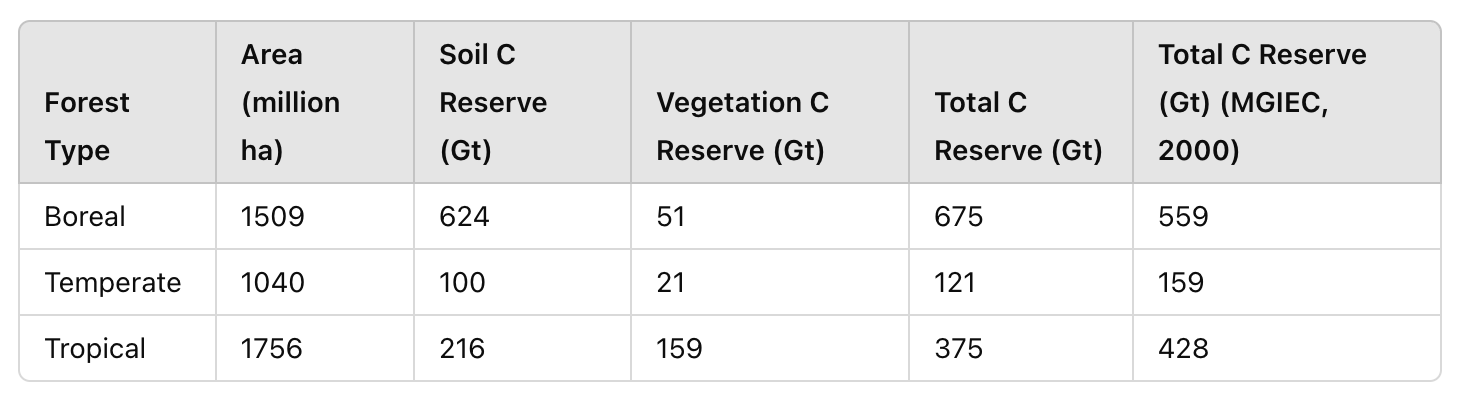
Most importantly, “global greening” is completely refuted by a fact precisely related to the “contours of destruction” (there are others, but this is a separate topic). Instead of the expected triggering of negative feedback (increased photosynthetic binding of CO2 and burial of C in the form of peat, removing carbon from the cycle — the main way of “cooling” the planet by biotic processes), alas, a positive connection is realized in the disturbed biosphere. An increase in CO2 concentration further accelerates the “eating” of organic matter previously buried in peat and other long-term carbon “storages”: litter, dead wood, litter, and soil (its volume is shown in Fig. 1). And this, in turn, accelerates the growth of CO2 concentration in the atmosphere: the circle closes.
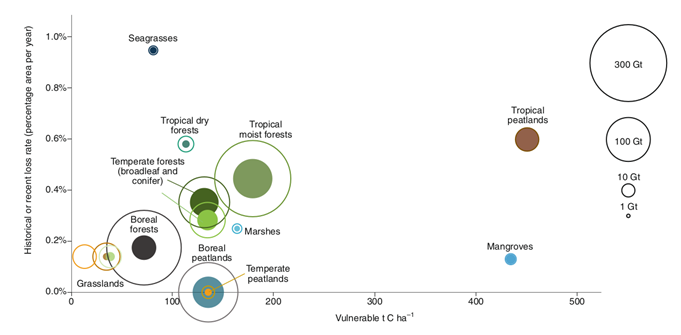
It turns out that ecosystems absorb an ever smaller share of the huge amount of carbon dioxide that enters the atmosphere each year due to the burning of fossil fuels, cement production, drainage of swamps, burning of vegetation, and other ecosystem disturbances. Until recently, the increase in CO2 emissions into the atmosphere was accompanied by greater binding of it by plants during photosynthesis, and to a lesser extent by ocean phytoplankton. However, since the 2000s. The growth of CO2 binding and C disposal has no longer “kept up” with the increase in its content in the atmosphere; since the 2010s it has completely begun to fall.
This was first discovered when studying the interannual shift in seasonal fluctuations in CO2 concentrations in the Northern Hemisphere. As global warming progresses, springs come earlier and earlier, photosynthesis begins earlier, so the binding of CO2 seems to dominate respiration more and more. However, the result contradicted expectations: climate changes in ecosystems stimulated respiration more strongly than photosynthesis, especially in the fall, so that as warming progresses, the dates in the fall, when respiration begins to prevail over photosynthesis, are coming earlier every year. Abnormally warm autumns, which occur more and more often, sharply increase the respiration of all organisms (not only plants but especially bacteria and fungi) according to Van’t Hoff’s rule. And this, the further, the more the annual season of predominance of CO2 binding over its release is shortened (instead of the expected lengthening), thereby negating the “increase” in C binding created by earlier springs.
“…The fact that the “carbon capacity” of the ocean has already reached its limit and the sequestration of additional amounts of CO2 by the ocean is decreasing has recently been proven by direct observations (Le Quéré C. et al., 2007). … an article was published in Nature on January 3, 2008, reporting alarming signs of declining CO2 absorption capacity also in terrestrial ecosystems. The authors … (16 people in total) focused their attention on the relationship between the long-term trend of increasing CO2 in the atmosphere and the regular seasonal fluctuations in CO2 concentration that occur against the background of this trend.
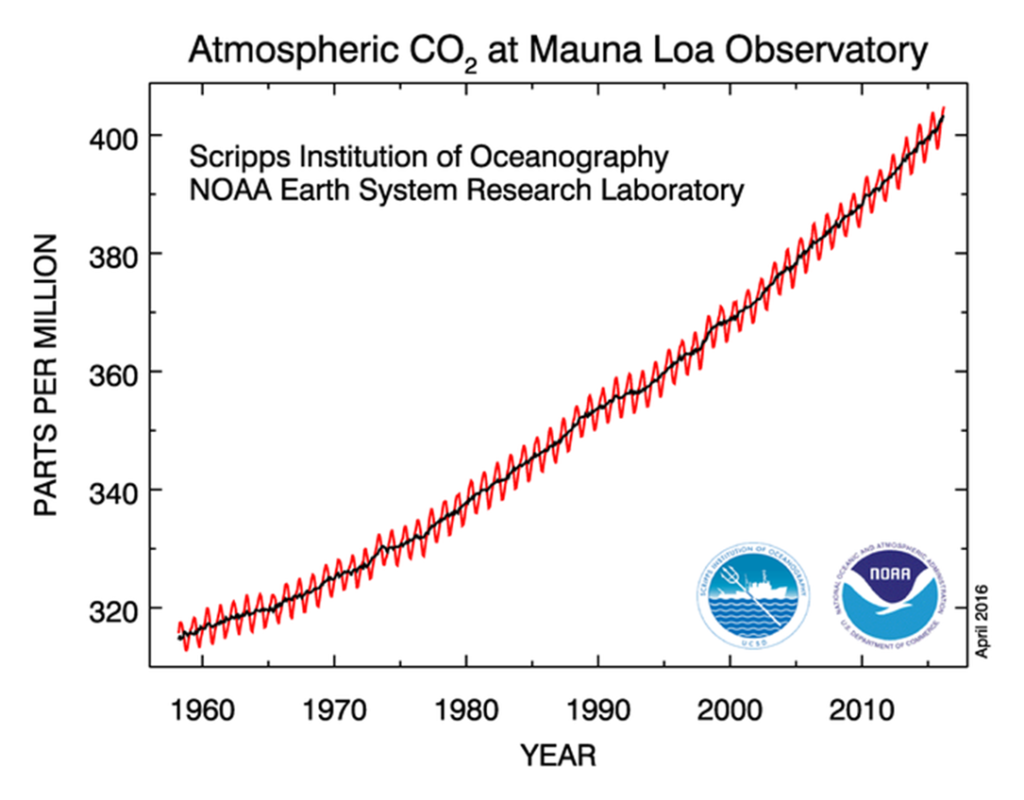
How these two dynamics (long-term and seasonal) relate can be illustrated using the example of the longest (almost half a century) series of data obtained by the Mauna Loa Observatory on the island of Hawaii. A graph summarizing the measurements shows both a continuous increase and small but regular seasonal variations in CO2 concentrations, with a maximum in April-May and a minimum in September-October. These fluctuations arise due to the fact that the process of consuming carbon dioxide, namely plant photosynthesis, occurs only during the warm period from late spring to late summer (in the northern hemisphere this is May-August). In autumn, winter and early spring, photosynthesis is impossible (at least in temperate and northern latitudes, where negative temperatures occur).
But in parallel, a process occurs in the ecosystem that is the opposite of photosynthesis — respiration (decomposition of organic matter with the consumption of oxygen and the release of CO2). Although all organisms breathe, the bulk of carbon dioxide entering the atmosphere is almost entirely determined by the respiration of decomposers, mainly bacteria and fungi. Respiration occurs over a longer period than photosynthesis. In summer, when it is warm, its intensity is especially high, but at the same time photosynthesis peaks, and as a result, much more CO2 is bound than is released. But once photosynthesis is weakened, the proportion of CO2 consumed and released shifts towards release, increasing the concentration of CO2 in the air.”
The authors of the work under discussion, using the example of several continuous (lasting at least 15 years) series of observations of changes in CO2 content in different points of the Northern Hemisphere, traced how the position of those points on the graph changes over time where the line of seasonal fluctuations intersects the line of the main trend. There are two such points per year: the “spring crossing” corresponds to the moment when the CO2 content curve goes down: as a result of intense photosynthesis, the processes of binding this gas begin to prevail over the release. The “autumn crossing” corresponds to the moment when the curve goes up and the release of CO2 as a result of respiration prevails over the binding (Fig. 2b).
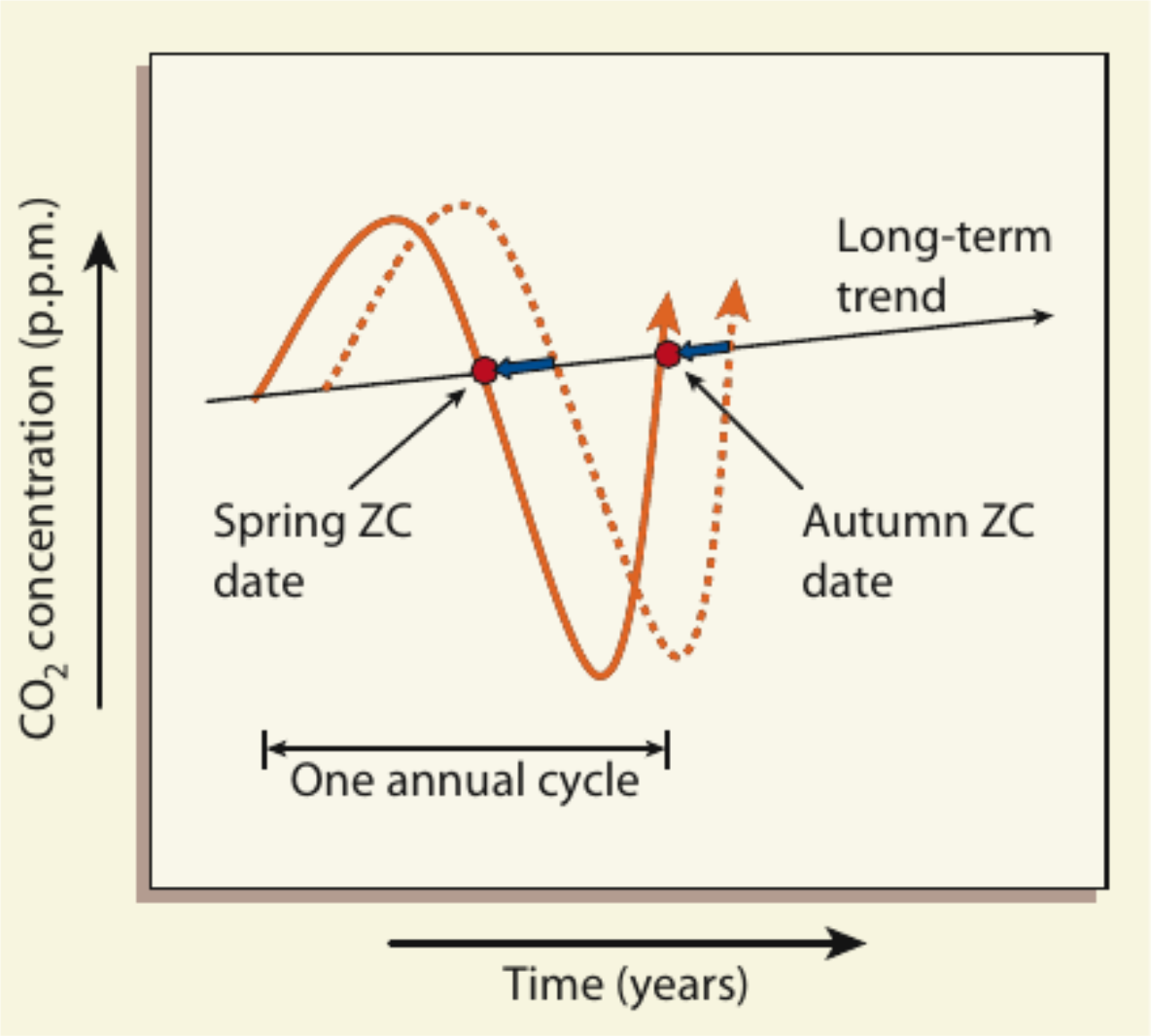
Until now, it was assumed that the increase in CO2 consumption by vegetation occurs primarily due to the lengthening of the growing season — the period of active plant growth. Indeed, spring is phenologically coming earlier (in Western Europe, compared to 1960, on average 12 days earlier), and autumn is increasingly abnormally warm. In theory, these climate changes (and the subsequent phenolate shift) will also affect the seasonal dynamics of CO2 content. The “spring crossing” should occur earlier and earlier, and the “autumn” crossing later (Fig. 2b).
In fact, the opposite happened: if the “spring crossing” actually began to be observed earlier, then the “autumn” one also moved to earlier dates and did not move to later dates, contrary to expectations. Thanks to this, the release of CO2 in the fall sharply increased, negating the increase in CO2 binding in a warmer and earlier spring (in the 2000s — by 90%, now — completely and even in excess). Piao et al. (2008) show that then the absorption of carbon by northern ecosystems (and not only) will noticeably decrease in the very near future, and the release will intensify and, the further, the more, it will overlap absorption.
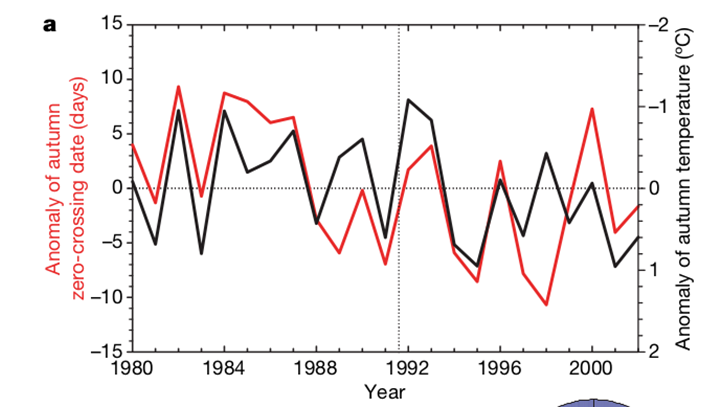
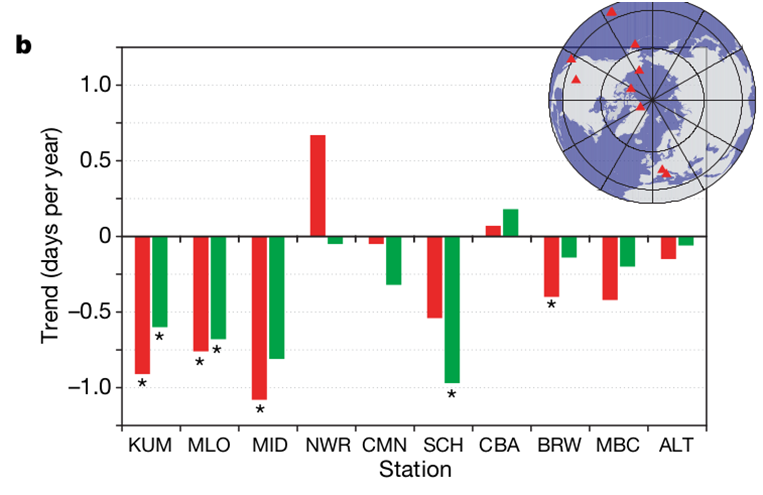
This sad forecast was more than justified, because these trends did not easily persist, but even intensified in the 2010s, spreading from the northern and temperate latitudes to the tropics (by about 2015). This is achieved primarily by shifting the “autumn crossing” to earlier and earlier dates, but not only. The first was recorded earliest — in the 1990–2000s. Piao et al. (2008) analyzed year-to-year changes in fall crossover dates, fall temperature anomalies, and carbon uptake predominant period (CUP) for 10 NOAA/ESRL network stations (air sampling for CO2 concentrations, CO2 fluxes in ecosystems, etc., over at least 15 years continuously). Their location in the Northern Hemisphere, see the red dots in the diagram in Fig. 3b. Also, using satellite data, we assessed the development of vegetation (NDVI index), gross (NEP), and net (NPP) production of ecosystems, as well as their respiration (TER).
Over all these years, the “autumn crossing” generally shifted to earlier dates (Fig. 3a), they negatively correlate with the anomalies of autumn temperatures around each station (in a wide band of ±20O latitude), for 4 out of 10 places the correlation is significant. The probability of obtaining such results purely by chance is 10–3, given the independence of records made at 10 stations.
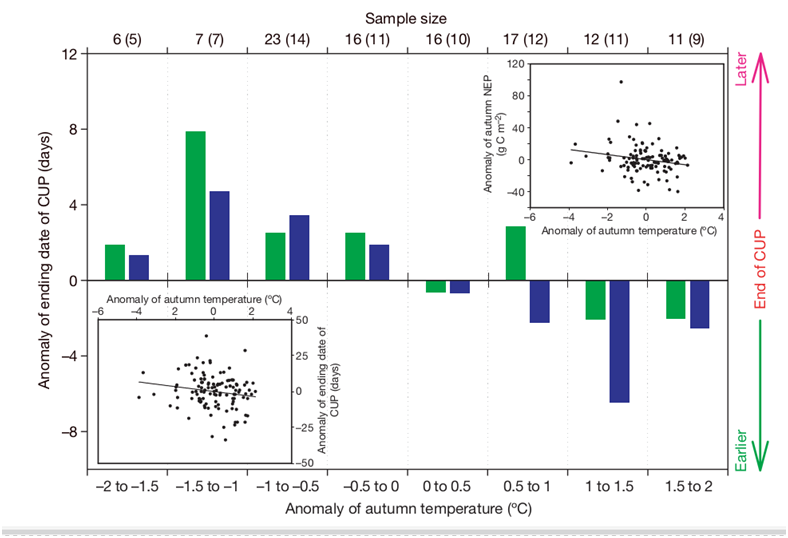
The authors refuted the assumption that the shift of the “autumn crossing” to earlier dates and the negative correlation of its value with autumn temperature anomalies are created by interannual differences in the intensity of winds that determine the transfer of CO2 in the atmosphere, establishing a strict connection with weather changes in the gross productivity of the NEP ecosystem: in warm autumns it accumulation ends earlier. They analyzed the influence of both factors in LMDZt, an atmospheric circulation model; With the exception of two observation sites (Mount Simone and Cape Kumukahi, CMN and KUM), the response was influenced specifically by changes in NEP, the effect of atmospheric transport was small. Increased ocean CO2 uptake and hydrocarbon combustion had little effect, explaining no more than 4% of the variance in the fall crossing date (excluding KUM). Changes in the seasonality of such burning, due to climate change, can only affect over time, and even then only slightly. This shift is not recorded in observations of the same stations for 1970–1994 but immediately appears later.
Simulating year-to-year changes in the productivity of terrestrial ecosystems in Northern Eurasia, depending on climatic variables and atmospheric CO2 transport (the ORCHIDEE model, “tuned” according to 1980–2002 data) refuted the competing explanation that the increase in autumn CO2 losses and the shortening of CUP is a consequence of greater the dryness of the summer preceding such an autumn. Weak summer minimums of CO2 absorption (a sign of drought) did not significantly affect these phenomena.
The shortening of CUP, recorded at almost all stations, and persisting in the future, means a decrease in the productivity of an increasing number of ecosystems over an expanding area, a reduction in phytomass instead of its accumulation, and the “burning” of carbon previously “stored” by the ecosystem instead of the expected increase in these stocks. This is exactly what is recorded when modeling the response of carbon flows through the response of ecosystems to global warming in ORCHIDEE, see Fig. 5, a — for changes in their gross photosynthetic productivity in autumn (GPP, more yellow and brown — decrease, greener and blue — increase, in Tg C/m2*OC), b — for changes in their net production in autumn, c — for changes in their gross production in autumn, d — for changes in NDVI.
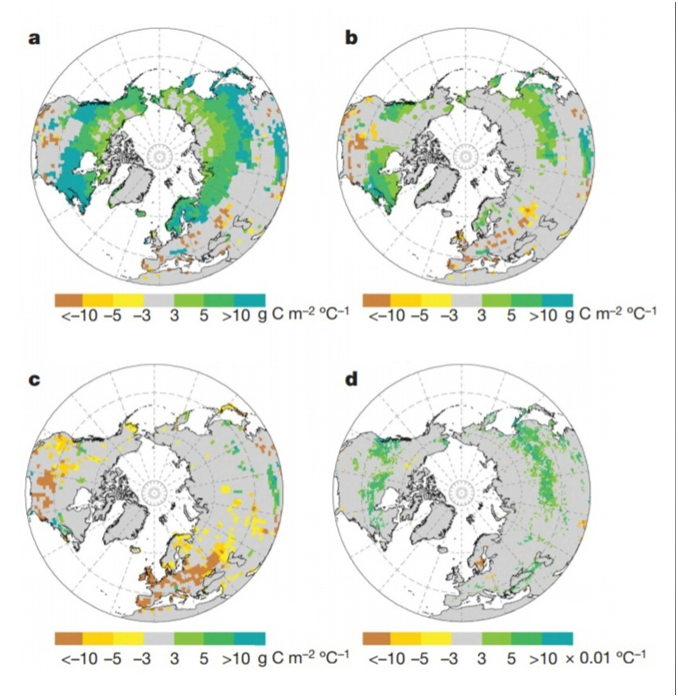
This effect, created by the “destruction circuit,” not only did not weaken in the following decades but sharply intensified and also spread from northern latitudes to tropical ones. Data on the forests of the northern hemisphere show a shift of the “spring crossing” forward by 1.9 ± 0.1 days but the autumn one by 2.6 ± 0.1 days, that is, CUP is reduced everywhere (each additional day of the growing season allows you to absorb more carbon, from 3 to 9.8 g/m2). Direct measurements of gas exchange at the leaf level indicate that this is associated with an earlier onset of leaf senescence and death during warm autumns (Zohner et al., 2023). All this is not counting the rise in the breath of decomposers in warm autumns!
Due to this and a number of other reasons, in 1980–2015. The photosynthetic response of ecosystems fell after a short rise, while respiration is constantly increasing, and by 2040–2060. in all terrestrial ecosystems, the release of CO2 will exceed absorption, instead of “global greening” there will be “browning”, global browing. By 2020, all biomes of the planet will respond to an increase in CO2 content by increasing its release, instead of the expected intensification of binding, that is, they will implement the classic destruction circuit.
Another beautiful destruction pattern has been described for desertification on the Mongolian Plateau (Peng Zhang et al., 2020). The authors analyzed the growth rings of coniferous trees over the past 260 years (they react especially sharply to abnormally hot and dry years, Fig. 7). In addition to the fact that they have been happening more often lately, each time they arrive triggers a positive feedback loop (Fig. 8). Soil moisture decreases and its temperature rises, which prolongs the heat period and makes its stressful effect on vegetation more and more stable (Fig. 6). The cover of vegetation, which was hitherto loose, is thinning out even more, the remaining plants are weakened, and the further this goes, the more difficult it is to recover, the process develops progressively.
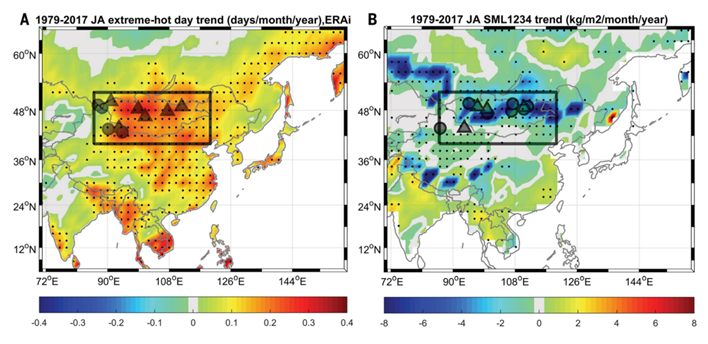
Let’s look at Figure 6. The black frame is the study area. The direction and amplitude of changes are shown in color; the more saturated the red, the more intense the increase in the parameter; the more saturated the blue, the more intense the decrease. Black dots are areas where identified trends are significant at P < 0.05: large triangles and circles are locations where tree-ring data was obtained and used to reconstruct intra- and interannual variations in the number of exceptionally hot days and soil moisture. At the same time, the water content of rivers is decreasing, the area of lakes is decreasing and their number is falling (by 26%) — as in general in this region, as well as in the Near and Middle East over the past 30 years. That is, thanks to this destruction circuit, the interaction of heat waves with overdried soil turns dry steppes and semi-deserts into deserts.
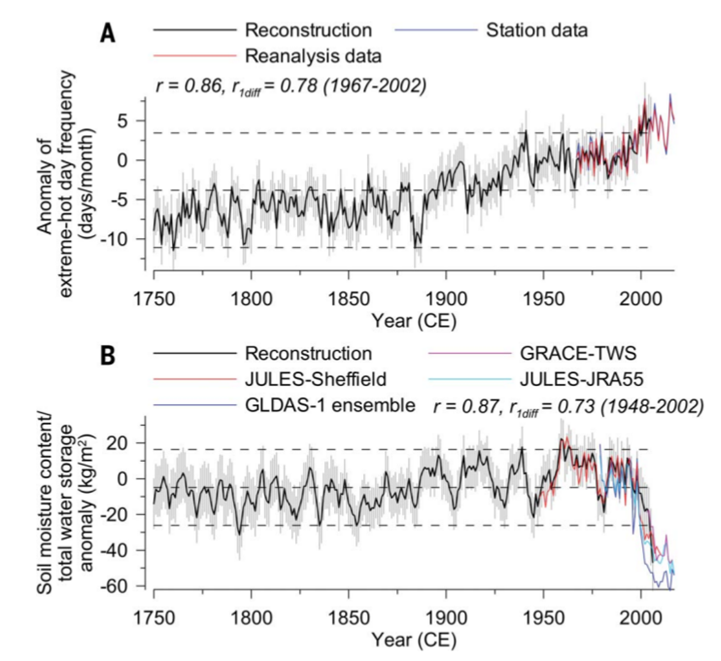
The launch of destruction contours creates a so-called tipping point, also known as the “point of no return”, when quantitative changes in the biogeocenosis under anthropogenic load turn into qualitative ones, irreversibly transforming it into something radically different (according to the existing typology of forests, habitats, landscapes and so on units). After the launch, reducing the impact alone does not solve the problem; you need to block the positive feedback that has turned on, and as quickly as possible. For example, desertification on the Mongolian plateau, having crossed a certain critical boundary, not only maintains itself but also intensifies it, see the film.
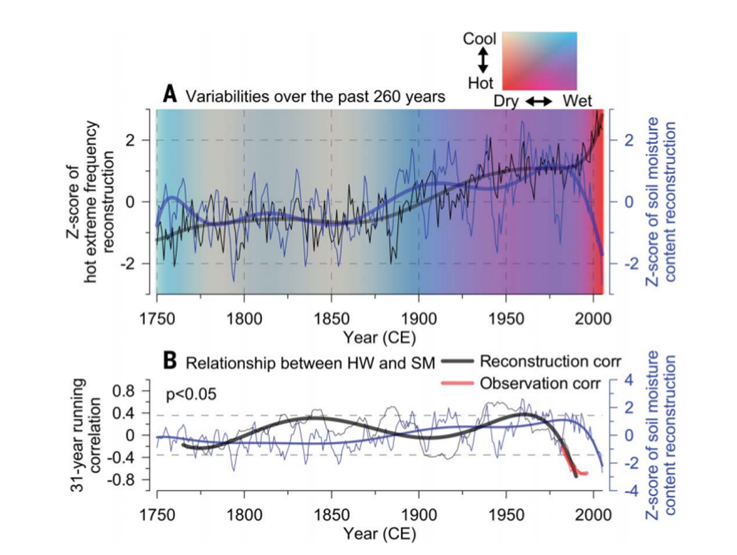
Let’s look at Figure 8. A. The actual variability of both parameters is shown by the values of the Z-criterion signs on the left abscissa for the first and right for the second (the larger Z, the more significant the variations in the (+) or (-) direction). Long-term trends of change are shown by the black and blue lines, respectively, as well as the saturation of the red and blue background colors: the redder, the hotter and drier, the blue — vice versa. B. Correlations of the number of extremely hot days (black lines) and soil moisture calculated over 31-year intervals based on tree-ring reconstruction data (1750–2005) or from the JULES-Sheffield project (1967–2011), light dotted line — correlations are significant at P<0.05. The blue line is long-term fluctuations in soil moisture, thin lines are low-frequency fluctuations of both parameters.
The third example of the launch of “circuits of destruction”: is the mass death of boxwood trees on the Black Sea coast of the Caucasus, which developed in the 2010s (including relict forests). The main reason for the environmental disaster is that during preparations for the Sochi Olympics, a phytophage, the boxwood moth Cydalima perspectalis, was brought in along with materials for landscaping; in the absence of predators and parasites, it multiplied and devoured everything. An important social point: quarantine authorities found a pest in one of the batches of planting material and demanded that it be destroyed. Gentlemen businessmen neglected this — profit is more expensive — and were not punished for this, and did not pay for the subsequent damage. And in the corresponding quarantine lists of the Russian Federation, either there is a fire, or there is not, and the law on such quarantine was adopted only in 2014.

However, the moth does not eat everything and everywhere; boxwood groves in gorges (the most shaded and wet places) might not be affected. Alas, they suffer most from another scourge — fungal diseases that lead to weakening and drying out of trees. And here a negative role was played, on the one hand, by long-term climate changes, and on the other, which is quite unexpected, by the epiphytic necker mosses Neckera crispa and neckera oblate mosses Neckera complanata, invariably present in boxwoods, covering the trunks and branches with a carpet.

Before climate change, the moss “sleeve” was useful for boxwood: during the hottest and driest days and weeks of summer, when the tree may suffer from drought, it reduced the corresponding stress because it accumulated and retained water, including through its condensation at night. Alas, the anthropogenic climate changes sweeping the planet have turned good into evil: the Russian subtropics have become noticeably warmer and much more humid, and much more precipitation falls there. In wetter summers, the moss “sleeve” already works against the tree, becoming a place of concentration of various pathogenic fungi that weaken the tree, and moisture increases vulnerability to them. Why are boxwoods not only “eaten”, but also “dry”: in addition, they are additionally weakened by the decline in snow cover in the low-mountain belt — also due to global warming.
“The processes of drying out were most clearly observed in the riverbed depressions of forest watercourses, characterized by more humid growing conditions. During the period from 2000 to 2011 in the Sochi region, the temperature increase was 0.5 OС, which was observed mainly in the warm period of the year. The strongest increase in temperature is observed in August. Of particular note is the increase in the number of days with temperatures above 25°C. In 2010 there were 59 such days, with an average of 20 days per year. For twenty-six days in 2010, the thermometer rose above 30 degrees, once above 36. In the foothill zone in Krasnaya Polyana during the study period, the average annual temperature ranged from 10.3OC (2004) to 13.4OC (2010), which exceeded the climatic norm at 0.8OC. The greatest changes relative to the climate norm were observed from July to October (more than +1.5OC).
According to meteorological observations from 2004 to 2010. on Aibga the temperature increased on average by 0.13OC per year, and on Dzhuta by 0.8OC. In 2010, the excess of the average long-term temperature at both observation points was more than 4OC. Since the early 1980s, Sochi has also experienced an increase in average monthly precipitation throughout the year (mainly in February-May and October). The amount of precipitation in February increased the most (by 40%). At the same time, the increase occurred due to an increase in days with high precipitation (more than 20 mm/day).
The greatest increase in precipitation is observed both in the foothill zone and at the high-mountain station “Dzhuga”, where the amount of precipitation in the cold period of the year almost doubled, and in the warm season — by 30%.
…According to the assessment of the forest pathological state of boxwood plantations, no trees without signs of weakening were noted. Larger trees of the main boxwood tier were generally in better condition. Natural regeneration is unsatisfactory. Undergrowth is sparse, withered and severely weakened plants predominate. Small plants were the most vulnerable. There was no self-seeding in most of the surveyed areas.
In each case of drying out, regardless of the type, there was a complex of pathogenic fungi that caused the death of thin branches and premature leaf fall. The dominant fungal species were Pseudonectria buxi (=Volutella buxi), Bionectria coronata (=Clonostachys buxi), as well as various species of Phomopsis and Macrophoma candollei.
In addition, for the first time in the plantings of the Sochi National Park, the causative agent of a dangerous boxwood disease, Cylindrocladium buxicola [an imported pathogen from Europe, which in Soviet times was steadily screened out by the quarantine service that had not yet collapsed]. However, based on the results of the analysis of numerous samples, it can be argued that its role in boxwood drying is not primary.
The species composition of pathogens, features of the manifestation of identified diseases, the nature of their spread, as well as a decrease in radial growth indicate that the development of pathogens is predetermined by the influence of other negative factors, including climatic ones.
…Thus, the root cause of the massive drying out and death of boxwood was the increase in summer temperatures over the last decade, as well as the seasonal redistribution of precipitation. These climatic changes could greatly affect the viability of individuals of the species Buxus colchica, as well as contribute to the massive spread and reproduction of pathogens… The current picture of the total destruction of boxwood trees by an invader — the boxwood moth — was a consequence of the importation of the species with planting material, in the absence of phytosanitary control and the failure to take urgent measures to destruction of introduced infected plants. In other words, the human factor was again decisive, which was preceded by a weakening of the viability of the ancient species.
The development of stress reactions in Colchian boxwood was facilitated by abnormal weather conditions of the last decade, manifested in an increase in temperature by 0.5°C, an increase in average monthly precipitation, and a decrease in snowfall in the low-mountain belt — the main area of growth of the species. All these climate changes have led to the weakening of Buxus colchica, and also contributed to the massive spread and proliferation of a complex of species of fungal pathogens that can cause premature leaf loss and weakening of plants, as well as the accumulation of high levels of infection.
An interesting feature of the damage and death of boxwood from pathogenic fungi was the presence of epiphytic mosses, primarily representatives of the genus Neckera, on the trunks and branches. Mosses greatly contributed to the preservation of the species, retaining moisture for a long time and creating a special microclimate for boxwoods. In the changing weather and climatic conditions of the last decade, mosses became a kind of “incubator” of pathogens and played a certain negative role when younger age groups and the lower parts of the trunks of tall trees, covered with a thick moss cover, were affected. This picture was observed primarily in stagnant areas of valley bottoms and side gorges protected from the wind, in microdepressions, while in blown areas of slopes and well-insulated biotopes, boxwood was more prosperous.”
What is the moral here? Two groups of factors (and the changes they create) are responsible for environmental problems everywhere and always: local and global, the latter in our case being anthropogenic climate warming and global trade in current export-oriented economies. If the local nature is disturbed and society is not in the best condition, they aggravate each other’s consequences; otherwise, they rather compensate. And if the contours of destruction are running, it is not enough to exclude the active anthropogenic factor — because of them, the destruction of ecosystems will continue with acceleration. They should be identified along with the influences that caused them and stopped with special measures.
illuminem Voices is a democratic space presenting the thoughts and opinions of leading Sustainability & Energy writers, their opinions do not necessarily represent those of illuminem.
illuminem briefings

Carbon Capture & Storage · Biodiversity
illuminem briefings

Carbon Capture & Storage · Biodiversity
illuminem briefings

Biodiversity · Nature
earth.com

Carbon Capture & Storage · Biodiversity
Grist

Carbon Capture & Storage · Biodiversity
CNN

Biodiversity · Nature